Предуведомление.
Как и прежде, публикуя материалы подобного рода, я не претендую на полную научность и корректность всех выводов. Я являюсь дипломированным историком, но не специалистом в области медицинских наук или генетической генеалогии. Нижеследующий очерк представляет собой попытку, на основе вспомогательных источников дополнить и расширить генеалогическое исследование истории моих предков. На этот раз по прямой восходящей женской линии.
Не скрою, я испытываю определенную гордость, ведь удалось последовательно выстроить линию из 9 поколений женщин, каждая из которых приходится последующей матерью. Полтора года назад мои знания обрывались на имени прапрабабушки – Наталье, о которой ровным счетом было ничего неизвестно. Теперь же, объединив работу с архивными источниками и данными генетиков мне есть, что рассказать читателям об этой ветви нашего рода.

Рис.1. Женщины нашей семьи (слева направо): моя мама Пепеляева Татьяна Васильевна (род. 1963), бабушка Пепеляева (Астраханцева) Раиса Григорьевна (род. 1940), прабабушка Астраханцева (Булатова) Татьяна Никифоровна (1904 - 1978 гг.)
***
Современное генеалогическое исследование, произведенное в отрыве от данных генетической генеалогии и генетики не может считаться целостным и завершенным. Письменные архивные источники официального (переписи и ревизии) и религиозного (метрические книги и исповедные росписи) характера, сколь полны бы они не были, не могут со 100% уверенностью установить факты супружеской неверности, случаи взятия приемного ребенка в семью, подмены ребенка или принятия новой женой детей мужа от предыдущих браков. Многие подобные моменты снимаются архивными источниками 2-й половины XIX века, но более ранняя эпоха оставляет большое пространство для маневра и рассуждений.
В своей прежней работе «ДНК-тестирование в генеалогических исследованиях: опыт практического применения» была предпринята попытка реконструкции истории моего рода, его дописьменного периода, на основе результатов ДНК-анализа, современных данных и тенденций в области изучения мужских Y-хромосомных гаплогрупп и гаплотипов. Но вопрос о митохондриальной ДНК, то есть происхождения по восходящей женской линии в этой статье не затрагивался и не рассматривался. Отчасти потому, что генезис мито-гаплогрупп, их вариативность и география распространения изучены сегодня не в пример менее подробно, чем у их Y-хромосомных партнеров. С другой стороны, сам процесс передачи женщиной своей ДНК более сложен и разнообразен, чем это происходит с наследованием мужской Y-хромосомы. Это связано с особенностями традиций брачности и социальных реалий минувших эпох. Так, девушек было принято выдавать в замужество в другие роды и племена, за несколько поколений их ДНК могла мигрировать на сотни километров и укорениться в совершенно новых общинах, в то время как их Y-хромосомные родичи могли продолжать жить на одном месте. Женщина также могла подвергнуться насилию со стороны соплеменников или завоевателей, быть захвачена в плен или продана – все её потомство, независимо от места и условий зачатия, получало материнскую мито-гаплогруппу.
В каждом поколении у мужчин мито-гаплогруппа меняется, а у женщин остается прежней. Частный вариант редукции предков предполагает «возвращение» пра-материнской мито-гаплогруппы потомкам одного Y-хромосомного рода. Такой случай может произойти, когда потомок одной пары родителей по мужской линии заключает брак с прямым потомком этой же пары предков по женской линии. В случае заключения такого брака, исходная мито-гаплогруппа будет присуща всем потомкам 1-го поколения такой пары, а среди дочерей – всех поколений, в которых родятся девочки. Проблема отслеживания подобного вида редукции обусловлена структурой письменных источников по генеалогии: если мужская линия, как правило, может быть надежно отслежена с конца XVI века, то аналогичное наблюдение женских линий становится возможным лишь с XVIII века, со многими оговорками.
Благодаря этим особенностям мито-гаплогруппы получают более широкое географическое распространение, имеют меньше выраженных очагов локализации и слабую степень ассоциации с национальным составом их носителей. Хотя некоторая корреляция между этносом и мито-гаплогруппами все же наблюдается, но не столь выражена, как аналогичное явление среди Y-хромосомных гаплогрупп.
Откуда в нашем геноме появилась митохондриальная ДНК и почему её так называют? На заре истории жизни на земле наши анаэробные предки-прокариоты «придумали» себе эволюционное преимущество, заключив симбиотический союз с органеллой под названием митохондрия. В результате в состав своего организма они инкорпорировали чужеродную форму жизни, обладавшую собственной ДНК. Первые получили эффективный способ питания с возможностью окисления органических соединений и использования освобождающейся при их распаде энергии. А вторые – надежную защиту, постоянный доступ к источнику питания и благоприятную среду для размножения. Союз оказался столь взаимовыгодным и прочным, что дал начало всем организмам на базе эукариотных клеток: фотосинтезирующих растений, грибов и животных, включая всех млекопитающих. Каждая клетка человека содержит от нескольких сотен, до нескольких тысяч митохондрий, однако на наследственность влияют только те, которые расположены в яйцеклетке. Большинство этих организмов, включая человека, способны наследовать митохондриальную ДНК только по женской линии, однако некоторые виды насекомых и даже млекопитающих могут унаследовать её по отцовской линии. Известны случаи, когда мито-ДНК передавалась потомкам именно от отца[1] из-за редкой генетической мутации, но в каждом таком случае это происходило путем включения ДНК митохондрий в ядерный геном. Правда таких случаев известно не много, они встречаются с частотой не более 0,06% и все они были обусловлены генетическими аномалиями.
Примечательно, что в последнее время ученые выявляют все больше случаев зависимости между наличием у человека определенной митохондриальной гаплогруппы или её снипа и вероятностью в будущем получить характерное заболевание. Например, заболевания меланомой среди лиц, у которых, как и у меня, в мито-ДНК присутствует SNP C16192T, встречаются в 1,7 раза чаще, чем его распространенность в популяции в целом[2]. В статье группы итальянских исследователей, под руководством Аурелии Санторо «Evidence for sub-haplogroup H5 of mitochondrial DNA as a risk factor for late onset Alzheimer's disease» утверждается, что люди с мито-гаплогруппой H5, особенно женщины, гораздо чаще (в 2 раза и более) подвержены риску заболевания болезнью Альцгеймера, и эта вероятность с возрастом только увеличивается[3].
Принимая во внимание вышесказанное, особенный практический интерес представляет собой составление мито-гаплогруппной базы жителей Вятки, до 1917 года, то есть до хронологической границы той эпохи, когда заключение брака между женихом и невестой одного вероисповедания, национальности, сословия и локации проживания перестало быть обязательным критерием и привело к быстрой и значительной миграции мито-гаплогрупп среди жителей этого региона.
Рискну предположить, что, как и в случае с Y-хромосомной популяцией жителей дореволюционной Вятки, мы не получим большого разнообразия вариантов мито-гаплогрупп, поскольку население региона было достаточно стабильным уже в сер. XVII века и подвержено, преимущественно, внутренней миграции, а также распространению из первоначальной ойкумены во вне. Также генетический состав популяции будет включать три характерных страты: мито-гаплогруппы коренного славянского населения северных уездов (Хлыновского, Орловского, Котельничского, Слободского, Шестаковского и Кайгородского), славянского населения южных уездов, переданных из состава Казанской губернии в 1780 гг. и мито-гаплогруппы финно-угорских народов: удмуртов, коми-пермян, коми-зырян, марийцев и др.
Выявление первого документированного носителя материнской мито-гаплогруппы потребует от исследователя обращения к изучению своей родословной по женским линиям, с обязательным установлением рода (семьи) из которого происходила женщина. Такое исследование, вероятно, будет ограничено известием 3-й ревизии 1762/63 гг. При этом, самые старшие женщины, упомянутые в указанной переписи, родились еще в конце XVII века, то есть в период, когда формирование этногенетического состава населения Вятки, в целом, уже завершилось. Указанные хронологические рамки позволяют проследить матрилинейную родословную ныне живущих людей на 9-11 поколений, не считая их собственного.
В моей семье прямая женская линия является «вятской», начиная от моей бабушки по материнской линии и далее вглубь веков, все женщины носители общей со мной мито-гаплогруппы проживали там.
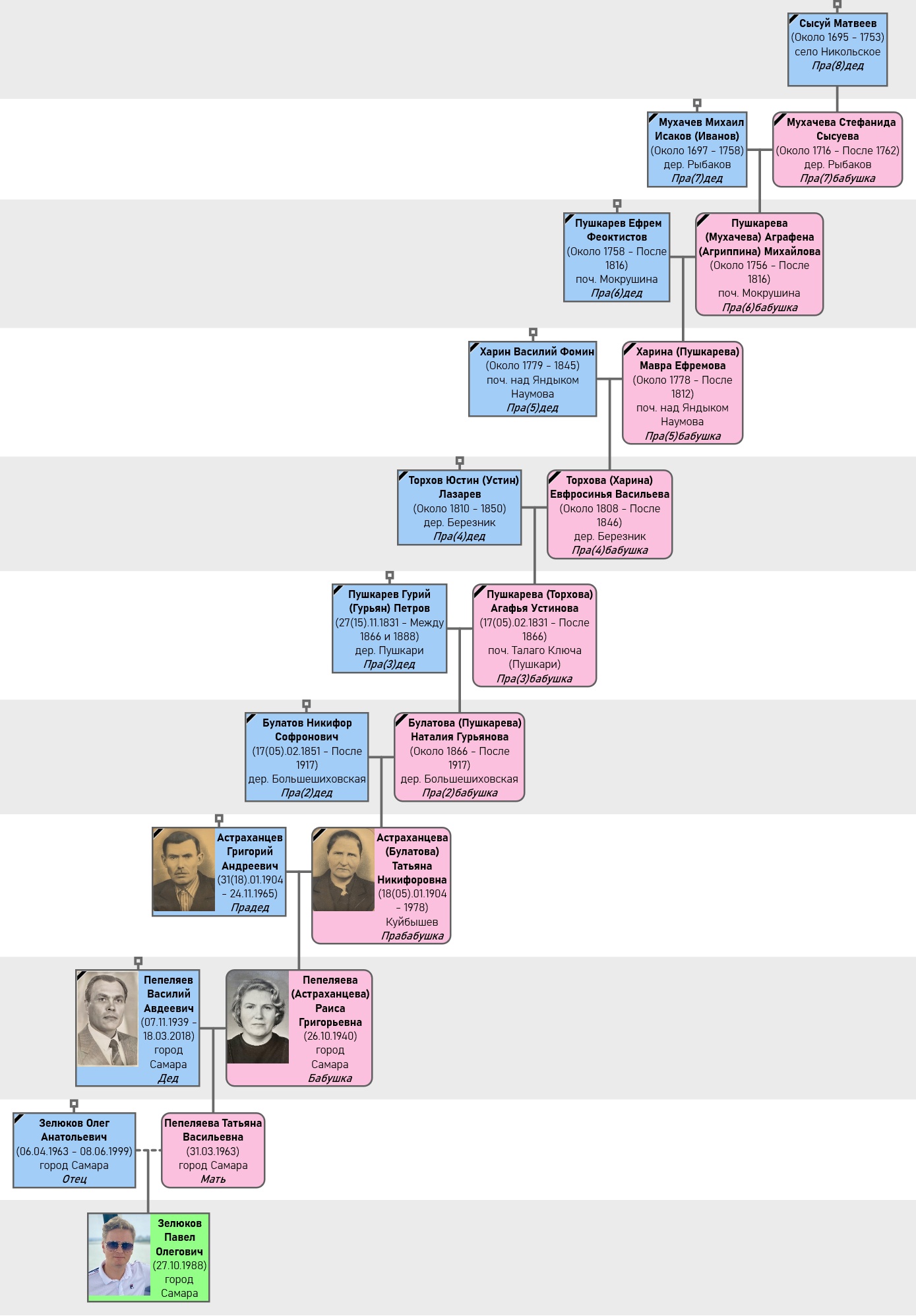
Рис. 2. Прямая восходящая линия моих предков по женской линии.
Если вести счет этой линии от моей матери, то получится следующая последовательность (в скобках указана девичья фамилия, место рождения):
- Пепеляева Татьяна Васильевна (род. 1963 г., мама);
- Пепеляева (Астраханцева) Раиса Григорьевна (род. 1940 г., бабушка, село Порез);
- Астраханцева (Булатова) Татьяна Никифоровна (1904 – 1978 гг., прабабушка, дер. Большое Шихово);
- Булатова (Пушкарева) Наталья Гурьева (ок. 1866 – после 1917 гг., прапрабабушка, дер. Пушкари);
- Пушкарева (Торхова) Агафья Устинова (1831 – после 1866 гг., пра (3 раза) бабушка, дер. Березник);
- Торхова (Харина) Евфросинья Васильева (ок. 1808 – после 1846 гг., пра (4 раза) бабушка, поч. над Яндыком Наумова);
- Харина (Пушкарева) Мавра Ефремова (ок. 1778 – после 1812 гг., пра (5 раз) бабушка, дер. Пушкари – поч. Талого Ключа);
- Пушкарева (Мухачева) Аграфена (Агриппина) Михайлова (ок. 1756 – после 1816 гг., пра (6 раз) бабушка, поч. Рыбаковский, дер. Рыбаки);
- Мухачева Стефанида Сысуева (ок. 1716 – после 1762 гг., пра (7 раз) бабушка, село Никольское Лобань).
Старейшим установленным предком по женской линии пока остается пра (7 раз) бабушка Мухачева Стефанида Сысуева (ок. 1716 – после 1762 гг.), которая родилась в селе Никольском (Лобань) у ясашного крестьянина Сысуя Матвеева (ок. 1695 – 1753 гг.), где он сам проживал с семейством еще на момент переписи 1710/11 гг. Фамилия рода самого Сысуя Матвеева достоверно не установлена – потомки его единственного сына Самсона (ок. 1728 – после 1762 гг.) покинули село Никольское до 4-й ревизии и их следы далее теряются. Учитывая, что само село Никольское (Лобань) было основано в 1701 году, Сысуй Матвеев, его отец Матвей Степанов (ок. 1640 – 1732 гг.) и мать Акулина Кондратьева (ок. 1642 – после 1719 гг.) были в числе его первопоселенцев.
Нужно отметить, что родной брат Сысуя Матвеева – Тимофей Матвеев (ок. 1675 – после 1720 гг.) служил священником в новопостроенном селе Ильинском (оно же Юртик на речке Ошланке) Уржумского уезда, о чем есть известие в ландратской переписи 1716 года (РГАДА. Ф. 350. Оп. 1. №146, Л. 942): «…в переписных книгах 1711 года написано у него Матвея сына Тимофея дочери Овдотья да Парасковья. И оной сын Тимофей до той переписи в попах в Уржумском уезде в селе Ошлане и вышеозначенные дочери его Тимофеевы и ныне у него…». Дальнейшее изучение этой ветки способно пролить свет на историю данного рода и открыть его исходную фамилию.

Рис. 3. Никольская церковь села Лобань на рисунке Татьяны Павловны Дедовой. Этот храм был местом, где десятки моих предков принимали крещение и венчались с конца XVIII до сер. XIX вв.
Имя матери Стефаниды Сысуевой пока остается для нас недоступно, так как эта женщина умерла до проведения ревизии 1762 года. Есть небольшой шанс найти исповедные росписи села Лобани за 1740-1750-е гг. и почерпнуть из них имя матери Степаниды, но пока это не сделано.
Поскольку нам известны имена, годы жизни, и место жительства всех моих предков по женской линии, их уместно наложить на карту. Мы видим, что в данном случае, женщины не проявляли высокой мобильности: за 250 лет и 8 поколений их гены «продвинулись» менее чем на 30 км от первоначального очага. Правда в каждом поколении женщины переселялись на новое место в дом мужа, «кочуя» по деревням и починкам Завятской новопоселенной, Лобанской и Порезской волостей. Дважды их домом становилась деревня Пушкари (поч. Талого Ключа): моя пра (3 раза) бабушка Торхова Агафья Устинова (1831 – после 1866 гг.) перешла туда жить к своему мужу Пушкареву Гурию Петрову (1831 – после 1866 гг.) из дер. Березник и там же родилась, и жила прежде, её собственная бабушка Харина (Пушкарева) Мавра Ефремова (ок. 1778 – после 1812 гг.).
Рис. 4. Маршрут "миграции" моих бабушек в XVIII - сер. XX вв.
Примечательно, что на моем поколении как раз обрывается эта линия мито-ДНК, включая всех потомков моей прапрабабушки Булатовой (Пушкаревой) Натальи Гурьевой (ок. 1866 – после 1917 гг.), так как все женские линии её потомков в настоящее время оставили только сыновей. Поэтому наши ближайшие генетические родственники по женской линии происходят либо от её сестры Маремьяны Гурьевой (род. 1856), либо от её теток Ксении (ок. 1827 – после 1858 гг.) и Авдотьи (ок. 1838 – после 1850 гг.) Устиновых Торховых.
Мито-гаплогруппой моей матери является H5-C16192T. Полные совпадения сочетания такого снипа и гаплогруппы, согласно базы данных проекта FamilyTreeDNA, в гипервариабельной области 1 (HVR1) выявлены еще у 9 участников проекта (3 в Польше, 2 в Финляндии, по 1 в Латвии, Литве, Швеции и Норвегии). В гипервариабельной области 2 (HVR2) с шестью персонами, представленными в первой группе (2 в Финляндии, по 1 в Польше, Литве и Норвегии. Полная последовательность с другими участниками, с дистанцией в 1 мутацию, выявлена с 3 лицами из первой группы (по одному в Польше, Литве и Норвегии).
Более внимательное изучение базы результатов проекта The Mitochondrial DNA Haplogroup H & HV Project позволило установить поименно 4 персоны с полным совпадением HVR1 и HVR2 маркеров: Анна Хенриксдоттер и Карин Хенриксдоттер из Финляндии, Кайса Эрсдоттер из Швеции – все жили в XVIII веке, и еще одна неназванная дама из современного Казахстана. Интересно, но не случайно, что тестирование аутосомной ДНК моей матери также выдает непропорционально большое количество «родственников» по женской линии из Швеции и Финляндии, а также обнаруживает присутствие 3% «финской» ДНК.
В отличие от ядерной ДНК, митохондриальная накапливает мутации примерно в 10 раз быстрее, так как она лишена защитных гистонов и её окружение чрезвычайно богато реактивными видами кислорода, являющимися побочным продуктом метаболических процессов, протекающих в митохондриях. Однако для передачи потомству, мутации должна подвергнуться именно митохондриальная ДНК органелл яйцеклетки, а не любой клетки вообще, поэтому для расхождения потомков на 1 мутацию должно потребоваться время, исчисляемое сроком жизни десятков поколений, вследствие чего наша общая с этими людьми «праматерь» могла жить как 300, так и 900 лет назад, но вряд ли ранее или точно не позже этого диапазона дат.
Что касается происхождения гаплогруппы H5, то традиционно её и родственные ей гаплогруппы, родоначальником которых выступает гаплогруппа H, возводят к представителям неолитических земледельческих культур Ближнего востока. Выделение H5 в самостоятельную гаплогруппу датируется временем около 13000 лет назад в Западной Азии. В последующем она с племенами земледельцев проникает в Европу, где, по данным археологических находок, получает широкое распространение в период между 6000 и 4000 лет назад. Масштабный анализ ДНК древних жителей Европы показал, что первые фермеры и последние охотники не враждовали друг с другом, а мирно сосуществовали и вступали в брак на протяжении многих тысяч лет[4].
Носители гаплогруппы H5 идентифицированы среди людей в захоронениях культуры литейно-ленточной керамики (5300-4900 гг. до н.э.), а также у представителей культуры боевых топоров (она же – культура шнуровой керамики, 2200-2000 гг. до н.э.). Во втором случае это может совпадать со временем проникновения указанной мито-гаплогруппы в племена, позже ставшие прото-славянами: венедами и антами. Еще один представитель гаплогуппы H5 был выявлен в неолитическом захоронении Мидхау на Оркнейских островах в Шотландии, с датировкой возраста 3630-3370 лет назад[5].
В статье группы польских ученых под руководством Алены Кушнаревич «Генетическое наследие балто-славяноязычных популяции: синтез аутосомных, митохондриальных и Y-хромосомных данных»[6] делается вывод о существовании матрилинейного единства восточных и западных славян (но не южных), а также непрерывности разнообразия мито-ДНК в их популяции, по крайней мере, на протяжении последних двух тысячелетий. При этом основу митохондриального разнообразия восточных и западных славян составили всего три гаплогруппы H5, H6 и U4a, которые были наиболее частыми в геномах жителей восточной Европы.
Российские ученые дополняют своих польских коллег. В совместной статье группы русско-польских ученых Института биологических проблем севера ДВО РАН и Института судебной медицины при университете Людвика Рюгдигера (Быдгощ, Польша), под руководством Б.А. Малярчука «Изменчивость митохондриальной ДНК у поляков и русских»[7] авторский коллектив предпринимает попытку выявить общепопуляционные закономерности распространения мито-гаплотипов и последовательностей SNP среди русских и поляков. Хотя авторы констатируют, что они не смогли найти каких-либо конкретных комбинаций уникальных гаплотипов мтДНК и их подкластеров, четко отличающих поляков и русских, как славяноязычные популяции, от соседних европейских популяций, таких как немцы и финны, они заметили некоторые закономерности частоты появления определенных подкластеров: «Другой H-подкластер, определяемый последовательностью мутаций 16189-16356 и его ответвления 16080-16189-16356 и 16189-16356-16362, часто встречался у поляков (3±3%), а также у русских (2±5%) и немцев (1±7%). Относительно высокая встречаемость H1-подкластера, определяемая последовательностью 16192-16304-16311 характерна для русских по сравнению с поляками и немцами, но другая H1-ветвь (с мотивом 16294-16304) явно является общим местом между немцами и поляками. Таким образом, Н5 в сочетании с последовательностью 16192-16304-16311 (у автора – отсутствует 16311) признается исследователями более характерным маркером русских, чем их соседей поляков и немцев.
Современное распространение гаплогруппы H5 среди жителей Европы, позволяет судить о её широкой географии в доиндоевропейский период истории. Области наибольшей концентрации (от 7 до 9% всей популяции) встречаются в регионах с автохтонным населением: Уэльсе, Альпийском регионе, Трансильвании, Латвии. Исключение представляет область вокруг современной Словении, куда H5 могла попасть как с племенами славян, так и быть воспринята от прежде населявших этот регион потомков представителей Кастельерской культуры, а также область Лацио вокруг Рима, где H5 могла быть унаследована от древнейшего населения Этрурии бронзового века – этрусков, признаваемого историками и археологами доиндоевропейским народом.
Субпопуляции из франко-кантабрийской области гаплогруппы H5 в сочетании с SNP 16192 (как у автора) могут быть коренными жителями Северной Испании, в частности, Астурии. Это очень гористый регион, исторически изолированный до XX века. Гаплогруппа H5 представлена у 1,86% астурийского населения и около 25% из них имеют рассматриваемую мутацию С16192Т. Этот факт служит надежным доказательством, что у этих людей и жителей Вятки XVI-XIX вв. существовали общие предки по женской линии, жившие по генетическим меркам совсем недавно.
Хронология древнейших носителей мито-гаплогруппы H5 в совокупности с SNP 16192-16304-16311 позволяет отследить пути миграции этих древних предков:
6800 – 6000 лет до н.э. – Сирия, Тель-Халула;
4625 – 4250 лет до н.э. – Германия, Обервидерштедт;
4400 – 3700 лет до н.э. – Греция, Караламбос;
2500 – 2050 лет до н.э. – Германия, Ротенширмбах;
800 г. до н.э. – 100 г. н.э. – Россия, Хакасия, Абакано-перевоз;
200 г. до н.э. – 100 г. н.э. – Польша, Гэски;
0 – 300 гг. н.э. – Польша, Ковалевско;
100 – 200 гг. н.э. – Польша, Рогово;
200 – 270 гг. н.э. – Дания, Сковгаарде;
900 – 950 гг. н.э. – Венгрия, Харта-Фрайфельт;
900 – 1100 гг. н.э. – Венгрия, Фад-Эгешеги;
1000 г. н.э. – Дания, о. Фюн;
1000 – 1400 гг. н.э. – Польша, Ледница;
XI век – Астрид (Эстрид) принцесса Датская, дочь Свена Вилобородого, мать Свена Эстридсена (родоначальника династии Эстридсенов);
1250 – 1450 гг. н.э. – Дания, Риисбю.
Приведенная хронология, далеко не полная, дает представление об этапах распространении H5-C16192T: 8-9 тыс. лет назад – локализация на Ближнем Востоке, 5,5-6,5 тыс. лет назад – проникновение в Европу широким фронтом от Германии до Греции, рубеж 1 тысячелетия до н.э. – 1 тысячелетия н.э. – укоренение в протославянской популяции антов. Вместе с тем, этот вариант мито-гаплогруппы укоренился также в популяциях кельтов и германцев, включая их британскую и скандинавскую ветви.
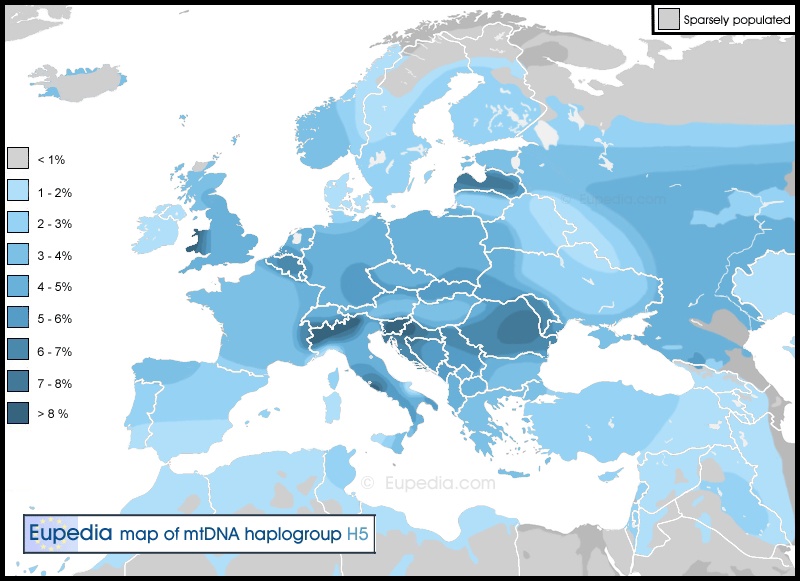
Рис. 5. Современное распространение носителей мито-гаплогруппы H5 среди населения Европы.
Ключевые отличия моей мито-ДНК от эталонной последовательности маркеров для гаплогруппы H5 – отсутствие C16311T и наличие C16519T (мутация, которую часто не принимают во внимание в силу высокой распространенности) в HVR1 и 309.1C, 309.2C, 315.1C, 522.1A, 522.2C, G3010A в HRV2. Все вместе они и определяют индивидуальность нашей мито-ДНК, но не дают ответа на вопрос, кем были в этническом плане мои далекие предки? Все, что мы можем констатировать с высокой долей уверенности, это то, что мои предки по женской линии проживали на Вятке ранее XVIII века и были русскими ясашными крестьянами, хотя пограничное (между Хлыновским, Казанским и Уржумским уездами) положение села Лобань открывает дорогу к большей вариативности рассуждений откуда именно прибыли первые женщины нашего рода.
Окончательный ответ на данный вопрос мы пока дать не можем, ведь документальная генеалогическая летопись не «пропускает» нас ранее первых десятилетий XVIII века. Остается надеяться, что база образцов ДНК будет пополняться и рано или поздно в ней появится больше совпадений, проливающих свет на древнюю историю наших далеких бабушек.
#история #генеалогия #ДНК #наследственность #род #предки #Вятка #семья #историясемьи #митохондрии
Очень серьезная проработка! Многим будет интересно ознакомиться с вашим трудом и подборкой материалов по распространению мито H5 на конкретном примере. Исследование непростое, требующее особой вьедливости и заинтересованности, а также времени. Мало кто так далеко сможет заглянуть, но у вас получилось! Администрация Родной Вятки наверное, уже подумывает о возможности создания специального раздела в блогах, выделяющих публикации на тему ДНК-генеалогии и связанные темы.😊 В новом году таких познавательных публикаций будет немало. 😊
Спасибо за отзыв! Я обеими руками «за», чтобы на Родной Вятке появился подобный раздел. Уверен, многие участники и гости портала также сдавали ДНК тесты, но делали это в разных лабораториях и единой базы нет. Сличение их результатов существенно обогатит нашу картину взаимосвязей вятский родов между собой.
Здравствуйте! Интересен внешний облик вашей прабабушки, в частности внешние углы глаз ( ближе к уху) - вытянутые, дающие длинные уклоняющиеся книзу линии тени. Я стал обращать внимание на такиеиглаза, но пока не нахожу зависимости - такой разрез у моего деда из под Орлова, у вашей прабабушки восточнее Нолинска, у маршала авиации Вершинина из юго-западной части Кировской, у Котельнических.
Здравствуйте! Интересную особенность вы подметили. Возможно есть какие то общие гены. Вершинины у нас в роду встречаются.
Здравствуйте! Моя бабушка, родившаяся в Орловском уезде, и известные мне ее мать и бабушка, также проживавшие там же, являются представителями гаплогруппы Н5 (Н5a1s по классификации YFull, в которой все - россиянки). Пересмотрел набор SNP, не нашел 16192 мутации. Если для вас будет интересно, готов поделиться результатами полногеномного теста (моего отца) по Мт.
Здравствуйте! Если нет снипа 16192, это означает, что генетическая дистанция достаточно большая, тысячелетия, если быть точнее.
Отлично вы понимаете. Для меня, к сожалению пока это китайская грамота.
мито Гаплогруппа из Вятских Полян очень редкая, азиатская D4E4а, в мутациях мы пока с дочкой и племянницей совсем не разобрались